English
КВАНТОВО-КЛАССИЧЕСКАЯ МОДЕЛЬ ПЕРЕНОСА ЭЛЕКТРОНОВ
Квантово-классическая модель моделирует перенос электрона вдоль цепочки, составленной из нескольких звеньев (называемых сайтами), каждый из которых представляется гармоническим осциллятором. В природе в роли такой цепочки служит двухцепочечная полимерная молекула ДНК, в которой мономеры - комплементарные нуклеотидные пары - связаны друг с другом посредством ковалентных связей. Нуклеотидные основания, образующие комплементарную пару, связаны между собой водородными связями. Модель учитывает взаимодействие между квантовой (заряд) и классической (нуклеотидные основания) системами. Молекула ДНК похожа на твёрдое тело тем, что пары оснований уложены в ней как в кристалле. Но это кристалл линейный, как бы одномерный - каждая пара оснований имеет только двух ближайших соседей. Основным отличием кристалла ДНК от обычных является его апериодичность, так как последовательность пар оснований в нём нерегулярна.
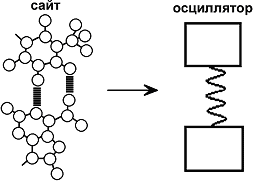 В исследуемой модели фрагмент ДНК рассматривается как цепочка, составленная из N сайтов. Каждый сайт представляет нуклеотидную пару, которая ведет себя как гармонический осциллятор. В роли пружины - водородная связь. В рассматриваемой модели считается, что плоскости оснований нуклеотидов параллельны друг другу, а расстояния между плоскостями оснований соседних сайтов постоянны, что соответствует B-форме ДНК. Молекула ДНК в равновесном состоянии не имеет свободных носителей заряда. Свободные электроны (анион-радикалы) или дырки (катион-радикалы) могут быть созданы либо посредством фотовозбуждения, либо в результате химических реакций.
В исследуемой модели фрагмент ДНК рассматривается как цепочка, составленная из N сайтов. Каждый сайт представляет нуклеотидную пару, которая ведет себя как гармонический осциллятор. В роли пружины - водородная связь. В рассматриваемой модели считается, что плоскости оснований нуклеотидов параллельны друг другу, а расстояния между плоскостями оснований соседних сайтов постоянны, что соответствует B-форме ДНК. Молекула ДНК в равновесном состоянии не имеет свободных носителей заряда. Свободные электроны (анион-радикалы) или дырки (катион-радикалы) могут быть созданы либо посредством фотовозбуждения, либо в результате химических реакций.
Перенос дырки в ДНК определяется перекрытием её волновых функций на соседних сайтах. Если перенос происходит вдоль одной из цепей ДНК, то интегралы перекрытия практически не зависят от внутрисайтовых смещений, независимо от величины последних. С учётом того, что масса нуклеотида на несколько порядков превышает массу дырки, смещения нуклеотидных пар можно описывать с помощью классических уравнений движения, а распространение дырки - квантово-механических. Гамильтониан переноса заряда вдоль цепочки сайтов имеет вид:
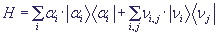 , (1)
, (1)
где αi - энергия электрона на i -ом сайте с волновой функцией  ,
,
νi,j - матричные элементы перехода с i -го на j -й сайт,  . Для описания переноса возбуждения будем искать решение уравнения Шредингера:
. Для описания переноса возбуждения будем искать решение уравнения Шредингера:
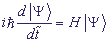 , (2)
, (2)
выбирая волновую функцию в виде 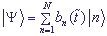 .
Величина bn представляет собой амплитуду вероятности нахождения электрона на n -ом сайте (
.
Величина bn представляет собой амплитуду вероятности нахождения электрона на n -ом сайте (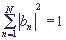 ).
).
Предполагаем, что энергия электрона на n -ом сайте является линейной функцией смещений сайтов 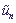 из их равновесных положений:
из их равновесных положений:
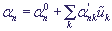 . (3)
. (3)
Подставляя выражения для H, Ψ и αn в уравнение Шредингера и учитывая, что νi,j = (νi,j)* = νj,i, i ≠ j (считая, что все коэффициенты - действительные числа), получаем:
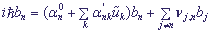 , n = 1,...,N, (4).
, n = 1,...,N, (4).
В отсутствии динамических флуктуаций (при α'nk = 0) система (4) описывает когерентное движение дырки. В общем случае движениями сайтов пренебрегать нельзя. В рассматриваемой модели мы предполагаем, что связь энергии дырки и смещений достаточно слабая, и учитываем в (3) только линейный член разложения.
Мы ограничились рассмотрением зависимости от смещений только диагональных матричных элементов αn в силу выбранной геометрии модели. Разумеется, в общем случае мы должны также учитывать и зависимость от смещений сайтов недиагональных матричных элементов ηn,j , которая может давать вклад такого же порядка. Более того, в ряде случаев именно вклад от смещений сайтов в недиагональную часть гамильтониана является основным.
Входящие в систему (4) смещения должны определяться из классических уравнений движения. Для получения этих уравнений введем кинетическую энергию сайтов  , Mk - масса k-го сайта, и потенциальную энергию
, Mk - масса k-го сайта, и потенциальную энергию 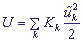 , Kk - упругая постоянная. Полный гамильтониан H рассматриваемой системы, усредненный по состоянию , имеет вид:
, Kk - упругая постоянная. Полный гамильтониан H рассматриваемой системы, усредненный по состоянию , имеет вид:
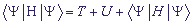 , (5).
, (5).
Объединяя уравнения (4) и уравнения движения для гамильтониана (5), получаем связанную систему уравнений, определяющую распределение заряда по N-сайтовой цепочке:
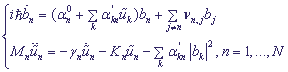 , (6).
, (6).
Здесь для учета процессов диссипации мы ввели член γn , γn - коэффициент трения.
Эта система является самосогласованной: эволюция амплитуд вероятностей bn определяется смещениями сайтов , которые, в свою очередь, зависят от распределения вероятностей |bn|2. Введение затухания в классическую подсистему (6) не является, вообще говоря, необходимым. Другой путь для моделирования подобных эффектов на конечных временах - введение большого числа классических степеней свободы для детализации модели, включая в рассмотрение отдельные атомы, входящие в состав ДНК, и молекулы окружающего ее растворителя.
В дальнейшем мы ограничиваемся случаем контактного взаимодействия: α'nk = δnkα'n, и предполагаем, что на динамику поведения сайта влияют только его ближайшие соседи, т.е. νi,j = 0, i ≠j ±1.Здесь δnk - символ Кронекера. Уравнения, соответствующие системе (6), имеют вид:
 , (7).
, (7).
Введённая таким образом модель является простейшей моделью, описывающей динамику переноса заряда в ДНК.
Соответствующая система в безразмерных переменных имеет вид:
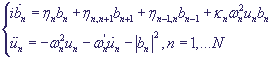 , (8).
, (8).
где τ = 10-14 - характерное время,
Un - характерный масштаб колебаний n -го сайта и
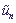 = Unun.
= Unun.
Потребуем выполнения соотношения α'nτ2/MnUn = 1, т.е.
Un = α'nτ2/Mn зависит от массы сайта и энергии электрона на сайте, а ωn - частота колебаний n -го сайта,
ηn = τα0n /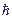 ,
ηn,n±1 = τνn,j /
,
ηn,n±1 = τνn,j / ,
ω2n = τ2Kn / Mn ,
ω'n = τγn / Mn ,
κn = α'nUnτ /.
,
ω2n = τ2Kn / Mn ,
ω'n = τγn / Mn ,
κn = α'nUnτ /.
Эффективная масса всех сайтов считается одинаковой и равной Mn = 10-21 г.
Высокочастотные внутримолекулярные колебания в ДНК, отвечающие колебаниям оснований в отдельном сайте,
имеют частоты порядка пикосекунд.
Частоты и коэффициенты трения на сайтах считаются одинаковыми; соответствующие безразмерные величины лежат в диапазоне:
ω2n ≈ 10-4 - 10-6, ω'n ≈ 10-2 - 10-5.
© В.Д.Лахно
Дополнительная информация
N.S.Fialko, V.D.Lakhno.
Long-range charge transfer in DNA.
Regular & Chaotic Dynamics, 2002, 7 (3)
V.D.Lakhno.
Sequence dependent hole evolution in DNA.
Journal of Biological Physics, 2004, 30, 123-138
�.�.�����, �.�.��������.
��������� � ������������� ��������� �������� ������ � ���.
���������, 2003, 48 (5), 797-801
| |